Description
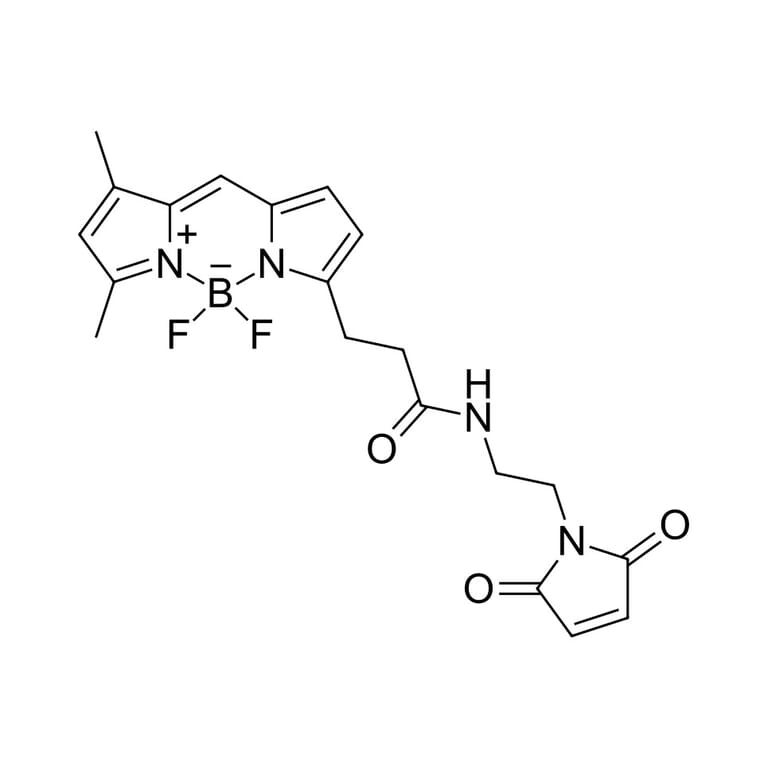
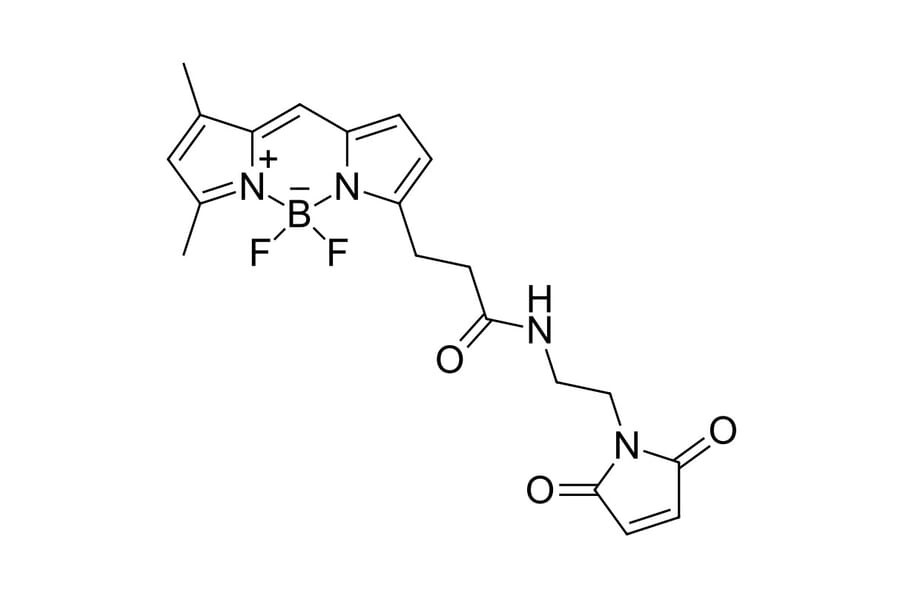
Thiol reactive BDP FL maleimide is a reactive dye for protein labeling, which has identical structure with BODIPY® FL maleimide. BDP FL is a borondipyrromethene dye which has absorption and fluorescence spectra similar to fluorescein (FAM). However, this dye exhibits very high photostability. It is non-charged, and has low molecular weight. Its brightness is similar to fluorescein, R110 and xanthene dye derivatives. This fluorophore is ideal for fluorescent microscopy and many other applications. The fluorophore can substitute fluorescein for almost any application, and it is compatible with any FAM-capable fluorescent instrumentation.