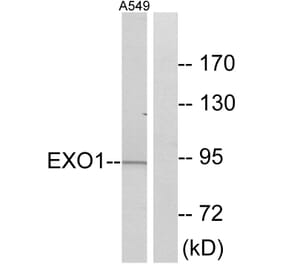
Exo1 (K86) pAb detects endogenous levels of Exo1 rotein.
Applications
WB, IF
Reactivity
Human, Mouse, Rat
Immunogen
Synthetic peptide, corresponding to amino acids 55-05 of Human Exo1.
Host
Rabbit
Clonality
Polyclonal
Conjugate
Unconjugated
Molecular Weight
~ 94 kDa
Purity
The antibody was affinity-purified from rabbit antiserum by affinity-chromatography using epitope-specific immunogen and the purity is > 95% (by SDS-PAGE).
Product Form
1 mg/ml in Phosphate buffered saline (PBS) with 15 mM sodium azide, approx. pH 7.2.
Synonyms
EXOI, Exonuclease 1, Exonuclease I, HEX1, hExo1, hExoI