Primary Antibodies
Secondary Antibodies
Proteins & Peptides
ELISA Kits
About Us
Contact Us
Sign In/Register
0
ISO 9001:2015 Certified
Live Customer Support
4.5/5 on Trustpilot
100% Quality Guarantee
Home
Primary Antibodies
Anti-Germ Cell-Specific Antigen Antibody (A423)
Anti-Germ Cell-Specific Antigen Antibody (A423)
Overview
Specifications
Images
Enlarge Image
Enlarge Image
Enlarge Image
$590
Product Datasheet
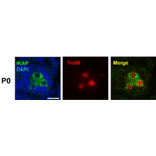
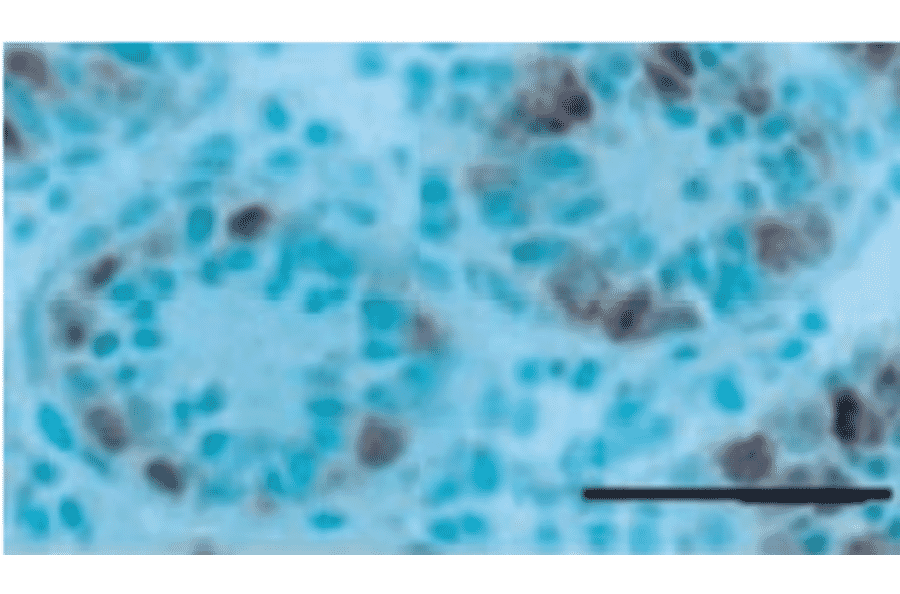
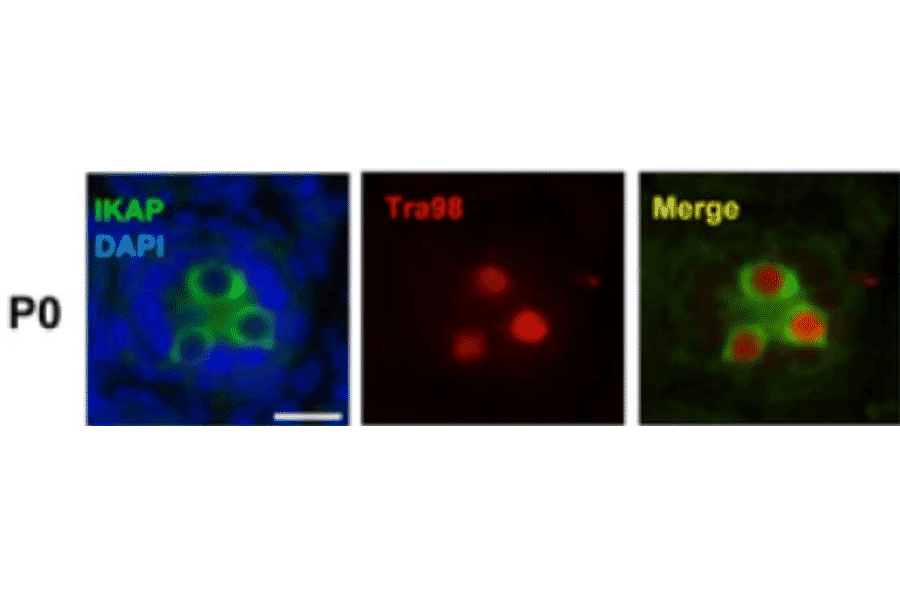
Rat monoclonal (TRA98) antibody to Germ Cell-Specific Antigen for WB, IP, IF, IHC-Fr and IHC-P.
100% Guarantee
Price Match Guarantee
Product Size:
100µg
Quantity:
1
2
3
4
5
6
7
8
9
10
Add To Cart
Request a Quotation
Custom or Bulk Request
Shipping Information
Freight/Packing Charges:
$40
Dispatched from St. Louis, MO.
Lead Time: 4-6 business days.
Specifications
Name
Anti-Germ Cell-Specific Antigen Antibody
Description
Rat monoclonal (TRA98) antibody to Germ Cell-Specific Antigen.
Applications
WB
,
IP
,
IF
,
IHC-Fr
,
IHC-P
Dilutions
WB: 1:1,000-1:5,000, IP: 1:200, IF: 1:400, IHC: 1:200-1:500
Reactivity
Mouse
Immunogen
Cell lysate of adult mouse testis.
Host
Rat
Clonality
Monoclonal
Clone ID
TRA98
Isotype
IgG2a
Conjugate
Unconjugated
Purification
Affinity purification.
Concentration
1 mg/ml
Product Form
Liquid
Formulation
Supplied in Phosphate Buffered Saline, pH 7.4, with 50% Glycerol (without Sodium Azide and carrier free).
Storage
Shipped at 4°C. Upon delivery aliquot and store at -20°C. Avoid freeze / thaw cycles.
Synonyms
GENA, Testicular germ cell-specific antigen
Isotype Controls
Rat IgG2a [RTG2A1-1] (A122131)
Rat IgG2a [RTG2A1-1] - BSA and Azide free (A122129)
Suitable Secondaries
Goat Anti-Rat IgG H&L Antibody (AP) (A294629)
Goat Anti-Rat IgG H&L Antibody (Biotin) (A294630)
Goat Anti-Rat IgG H&L Antibody (FITC) (A294635)
Goat Anti-Rat IgG H&L Antibody (HRP) (A294636)
See all Anti-Rat IgG Secondaries →
Disclaimer
This product is for research use only. It is not intended for diagnostic or therapeutic use.
Scientific Validation Data
Validation Data
(3)
Enlarge Image
Anti-Germ Cell-Specific Antigen Antibody (A423)
Anti-Germ Cell-Specific Antigen Antibody
Enlarge Image
Anti-Germ Cell-Specific Antigen Antibody (A423)
Anti-Germ Cell-Specific Antigen Antibody
Enlarge Image
Anti-Germ Cell-Specific Antigen Antibody (A423)
Anti-Germ Cell-Specific Antigen Antibody
Publishing research using Anti-Germ Cell-Specific Antigen Antibody (A423)? Please
let us know
so that we can list the citation on this page.
Top