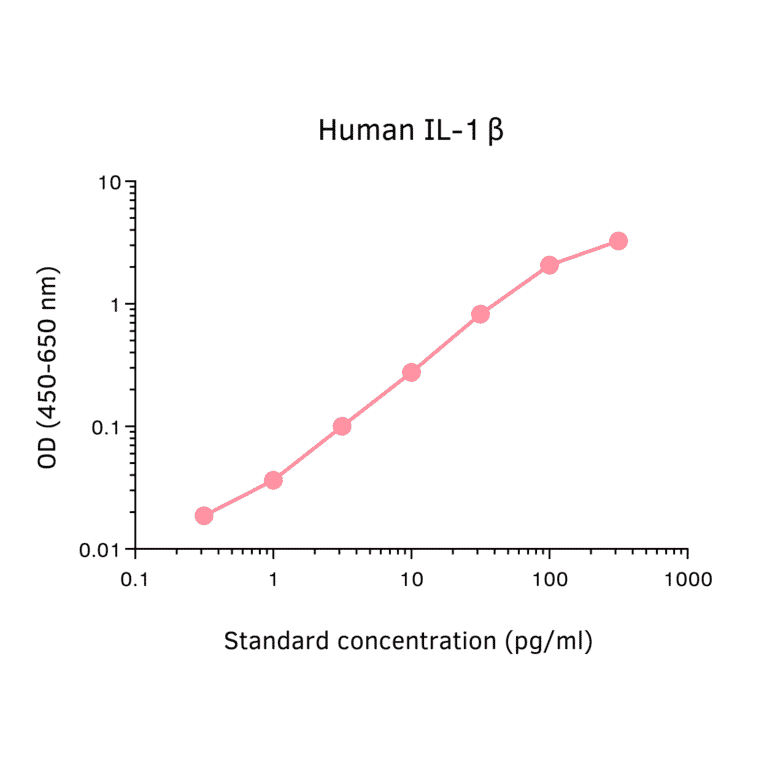
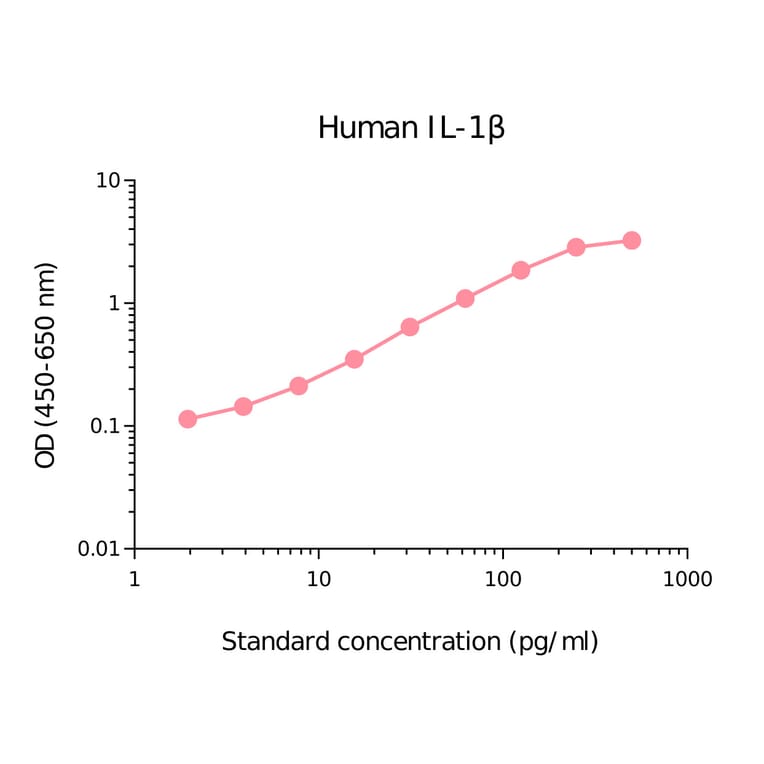
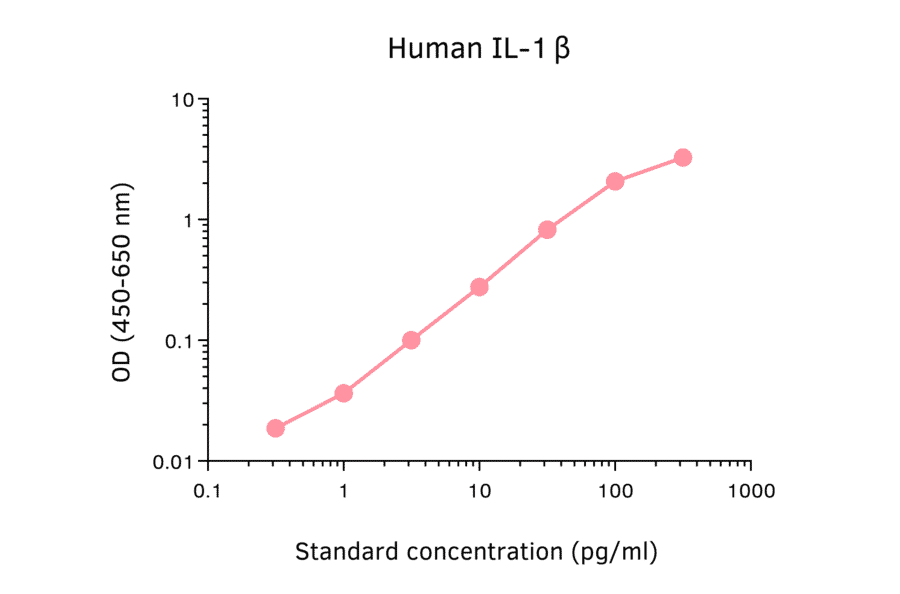
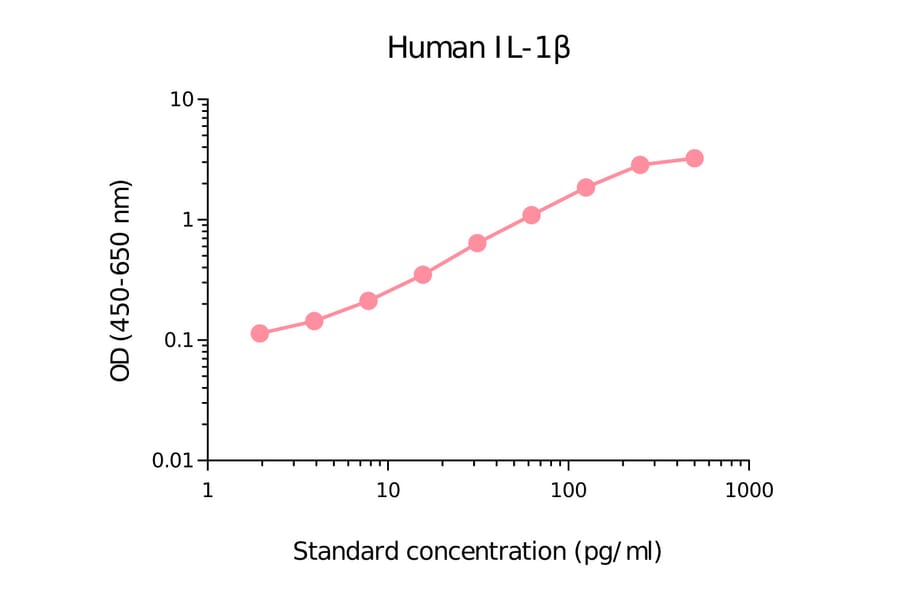
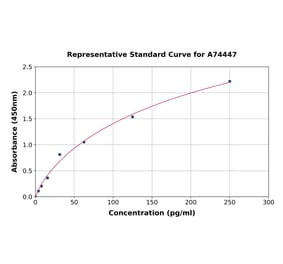
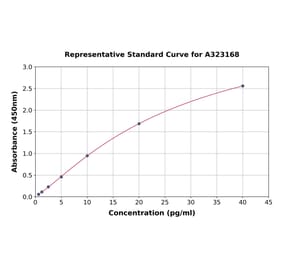
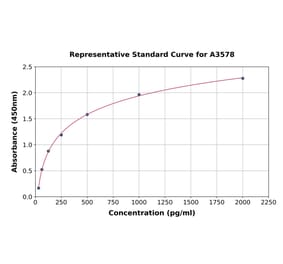
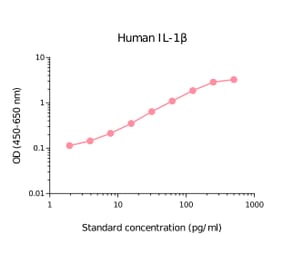
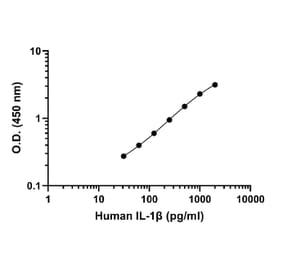
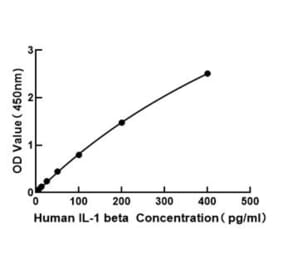
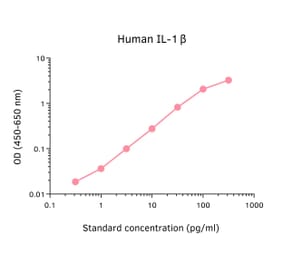
Human IL-1 beta ELISA Kit is a sandwich Enzyme-Linked Immunosorbent Assay (sELISA) designed for the in vitro quantitative determination of Human IL-1 beta in serum, plasma, and cell culture supernatant.
Assay Type
Sandwich (quantitative)
Platform
Pre-coated Microplate (12 x 8 well strips)
Detection Type
Colorimetric
Instrument
Colorimetric Microplate Reader
Sample Type
Serum, plasma, and cell culture supernatant.
Sensitivity
0.4 pg/ml
Product Range
1-316 pg/ml
Specificity
This ELISA kit utilizes a pair of antibodies to specifically recognize human IL-1 beta (pro-recognizem and active recognizem).
Reactivity
Human
Cross Reactivity
This ELISA antibody does not cross react with human IL-1 alpha.
Storage
Shipped at +4°C. Store the ELISA standard at -20°C. Store all other reagents at +4°C.
Synonyms
Catabolin, IL1B, IL1F2, Interleukin-1 beta
Product Note
Information online is representative. Always refer to the Product Manual inside the kit.
Disclaimer
This product is for research use only. It is not intended for diagnostic or therapeutic use.