Primary Antibodies
Secondary Antibodies
Proteins & Peptides
ELISA Kits
About Us
Contact Us
Sign In/Register
0
ISO 9001:2015 Certified
Live Customer Support
4.5/5 on Trustpilot
100% Quality Guarantee
Home
Proteins and Peptides
PCNA Proteins
Recombinant Human PCNA Protein (Functional) (A476)
Recombinant Human PCNA Protein (Functional) (A476)
Overview
Specifications
Images
Enlarge Image
Enlarge Image
$375
Product Datasheet
100% Guarantee
Price Match Guarantee
Product Size:
20µg
100µg
Quantity:
1
2
3
4
5
6
7
8
9
10
Add To Cart
Request a Quotation
Custom or Bulk Request
Shipping Information
Freight/Packing Charges:
$40
Dispatched from St. Louis, MO.
Lead Time: 4-6 business days.
Tags:
Biologically Active
Specifications
Name
Recombinant Human PCNA Protein (Functional)
Applications
WB, DB, SDS-PAGE, ELISA, Functional Studies
Expression System
Escherichia coli
Nature
Recombinant
Protein Species
Human
Protein Length
Full length protein.
Molecular Weight
29 kDa
Biologically Active
Yes
Purity
> 98% (by SDS-PAGE).
Product Form
Liquid
Concentration
1.0 mg/ml
Formulation
Supplied in 25 mM HEPES, pH 7.9, with 1 mM EDTA, 0.01% NP40, 1 mM DTT, 2 µg/ml Leupeptin, 0.1 mM PMSF, 75 mM NaCl, and 50% Glycerol.
Storage
Shipped at 4°C. Upon delivery aliquot and store at -20°C. Avoid freeze / thaw cycles.
Synonyms
Cyclin, Proliferating cell nuclear antigen
Disclaimer
This product is for research use only. It is not intended for diagnostic or therapeutic use.
Scientific Validation Data
Validation Data
(2)
Enlarge Image
SDS-PAGE - Recombinant Human PCNA Protein (Functional) (A476)
Recombinant Human PCNA Protein (Functional)
Enlarge Image
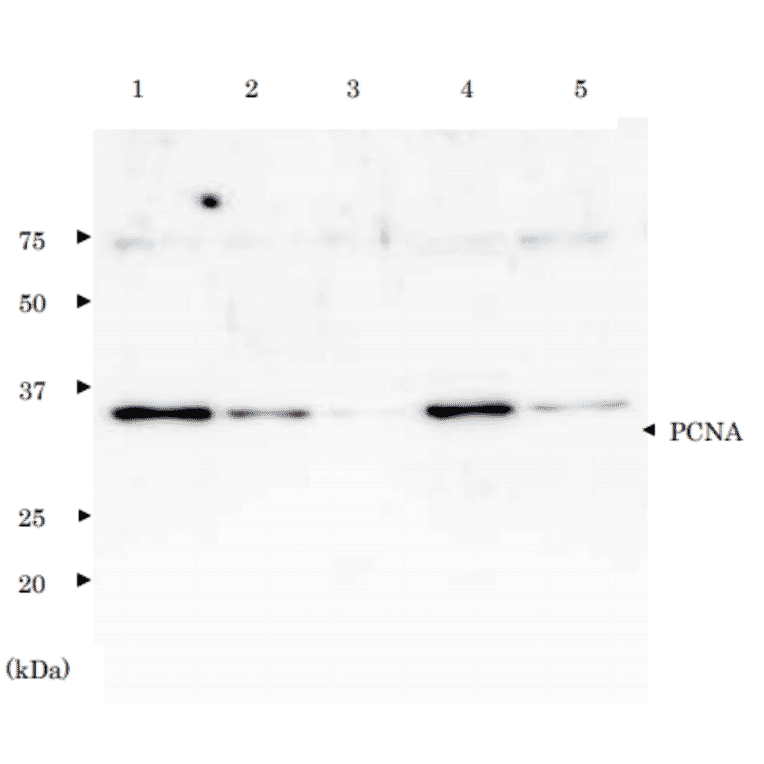
Western Blot - Recombinant Human PCNA Protein (Functional) (A476)
Recombinant Human PCNA Protein (Functional)
Publishing research using Recombinant Human PCNA Protein (Functional) (A476)? Please
let us know
so that we can list the citation on this page.
Alternative products to Recombinant Human PCNA Protein (Functional) (A476)
A57804
Recombinant Human PCNA Protein (His Tag)
Validated for WB, ELISA.
A60829
Recombinant Human PCNA Protein
Validated for SDS-PAGE.
A57804
Recombinant Human PCNA Protein (His Tag)
Validated for WB, ELISA.
A60829
Recombinant Human PCNA Protein
Validated for SDS-PAGE.
Proteins predicted to interact with PCNA
Predicted protein interactions based upon String database. Revelancy score correlates with probability of interaction.
DNA polymerase eta Proteins
99.9% Relevancy Score
FEN1 Proteins
99.9% Relevancy Score
p21 Proteins
99.9% Relevancy Score
MSH6 Proteins
99.8% Relevancy Score
KIAA0101 Proteins
99.7% Relevancy Score
CDK4 Proteins
99.6% Relevancy Score
POLD4 Proteins
99% Relevancy Score
Cyclin D1 Proteins
98.9% Relevancy Score
GADD45A Proteins
98.7% Relevancy Score
Top