The c-KIT receptor tyrosine kinase mediates the cellular response to stem cell factor (SCF). Whereas c-KIT activity is important for the proliferation of hematopoietic cells, melanocytes and germ cells, uncontrolled c-KIT activity contributes to the growth of diverse human tumors. Suppressor of cytokine signaling 6 (SOCS6) is a member of the SOCS family of E3 ubiquitin ligases that can interact with c-KIT and suppress c-KIT-dependent pathways. Here, we analyzed the molecular mechanisms that determine SOCS6 substrate recognition. Our results show that the SH2 domain of SOCS6 is essential for its interaction with c-KIT pY568. The 1.45-Å crystal structure of SOCS6 SH2 domain bound to the c-KIT substrate peptide (c-KIT residues 564-574) revealed a highly complementary and specific interface giving rise to a high affinity interaction (K(d) = 0.3 μm). Interestingly, the SH2 binding pocket extends to substrate residue position pY+6 and envelopes the c-KIT phosphopeptide with a large BG loop insertion that contributes significantly to substrate interaction. We demonstrate that SOCS6 has ubiquitin ligase activity toward c-KIT and regulates c-KIT protein turnover in cells. Our data support a role of SOCS6 as a feedback inhibitor of SCF-dependent signaling and provides molecular data to account for target specificity within the SOCS family of ubiquitin ligases.
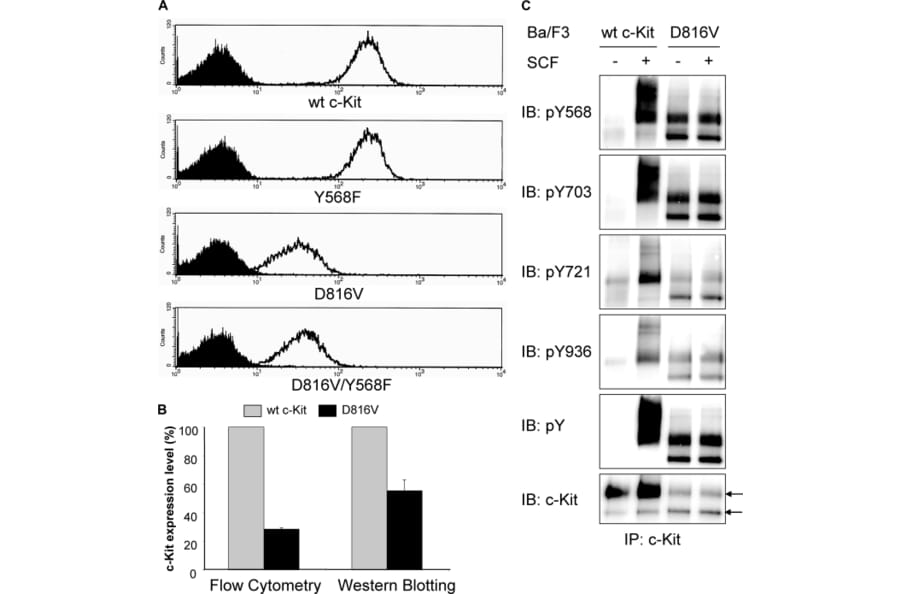
The receptor tyrosine kinase c-Kit plays a critical role in hematopoiesis, and gain-of-function mutations of the receptor are frequently seen in several malignancies, including acute myeloid leukemia, gastrointestinal stromal tumors, and testicular carcinoma. The most common mutation of c-Kit in these disorders is a substitution of the aspartic acid residue in position 816 to a valine (D816V), leading to constitutive activation of the receptor. In this study, we aimed to investigate the role of Src family kinases in c-Kit/D816V signaling. Src family kinases are necessary for the phosphorylation of wild-type c-Kit as well as of activation of downstream signaling pathways including receptor ubiquitination and the Ras/Mek/Erk pathway. Our data demonstrate that, unlike wild-type c-Kit, the phosphorylation of c-Kit/D816V is not dependent on Src family kinases. In addition, we found that neither receptor ubiquitination nor Erk activation by c-Kit/D816V required activation of Src family kinases. In vitro kinase assay using synthetic peptides revealed that c-Kit/D816V had an altered substrate specificity resembling Src and Abl tyrosine kinases. We further present evidence that, in contrast to wild-type c-Kit, Src family kinases are dispensable for c-Kit/D816V cell survival, proliferation, and colony formation. Taken together, we demonstrate that the signal transduction pathways mediated by c-Kit/D816V are markedly different from those activated by wild-type c-Kit and that altered substrate specificity of c-Kit circumvents a need for Src family kinases in signaling of growth and survival, thereby contributing to the transforming potential of c-Kit/D816V.