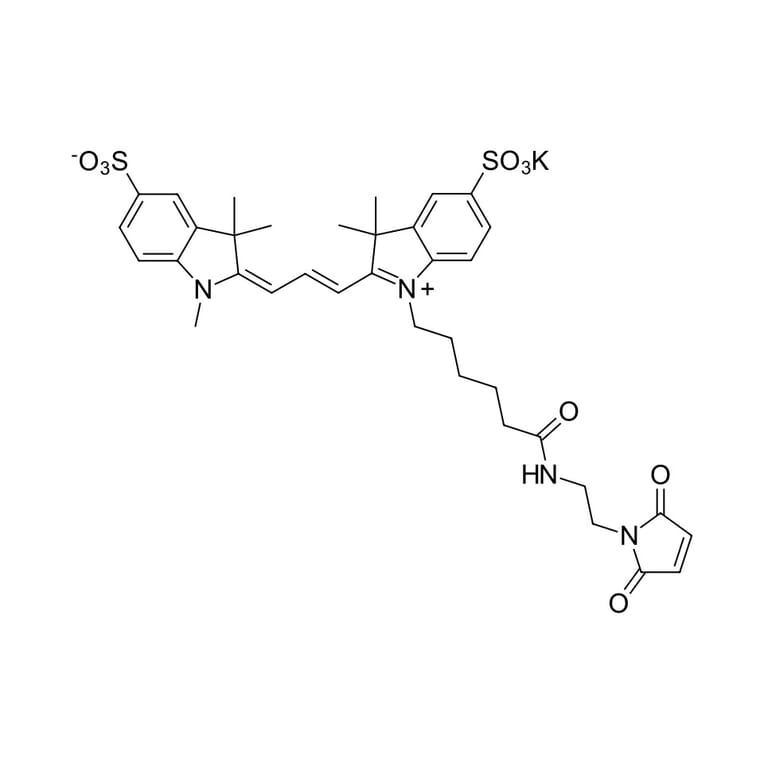
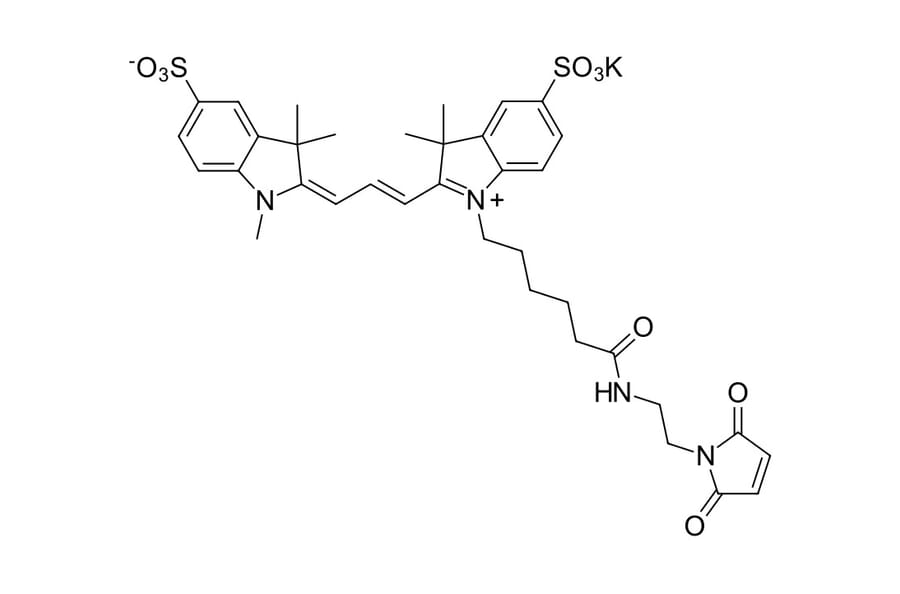
Sulfo-Cyanine 3 maleimide is a water soluble, thiol reactive dye for the labeling with hydrophilic sulfo-Cyanine 3 fluorophore. This is an analog of Cy3® maleimide. This product is recommended for the labeling of antibodies and other labile proteins in mild, purely aqueous conditions. The dye is water soluble and does not require use of organic co-solvent. A non-sulfonated Cyanine 3 maleimide is available.
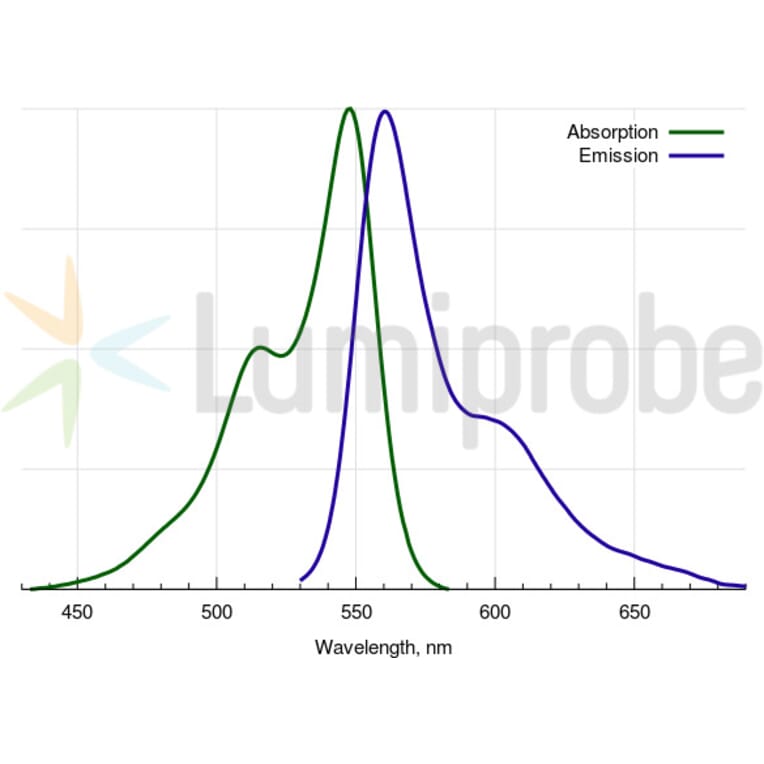
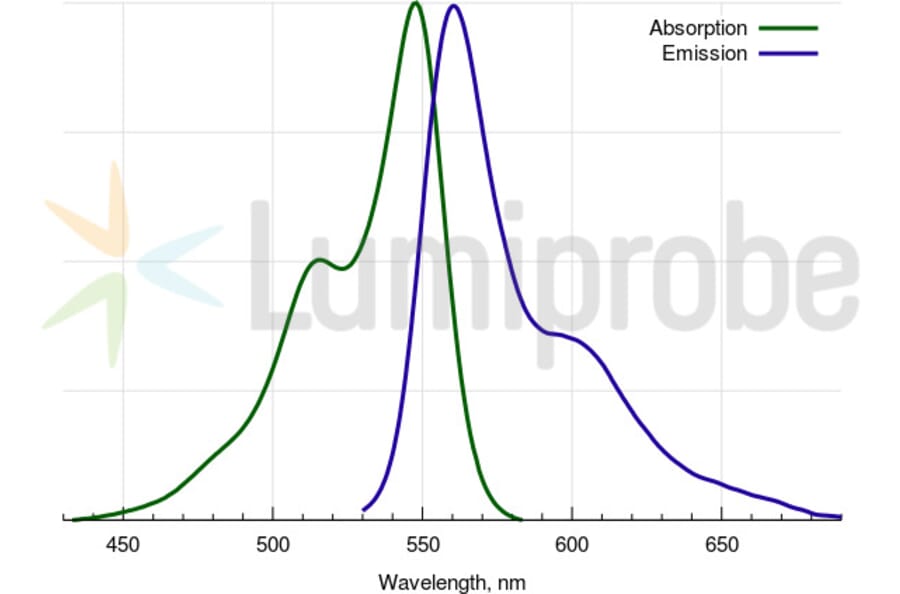
Absorption Maxima
548 nm
Extinction Coefficient
162000 M-1cm-1
Emission Maxima
563 nm
Fluorescence Quantum Yield
0.1
CAS Number
1656990-68-9
CF260
0.03
CF280
0.06
Purity
> 95% (by 1H NMR and HPLC-MS).
Molecular Formula
C36H41KN4O9S2
Molecular Weight
776.96 Da
Product Form
Red powder.
Solubility
Soluble in water (0.39 M = 30 g/L), DMF, and DMSO.
Storage
Shipped at room temperature. Upon delivery, store in the dark at -20°C. Avoid prolonged exposure to light. Desiccate.
Disclaimer
This product is for research use only. It is not intended for diagnostic or therapeutic use.