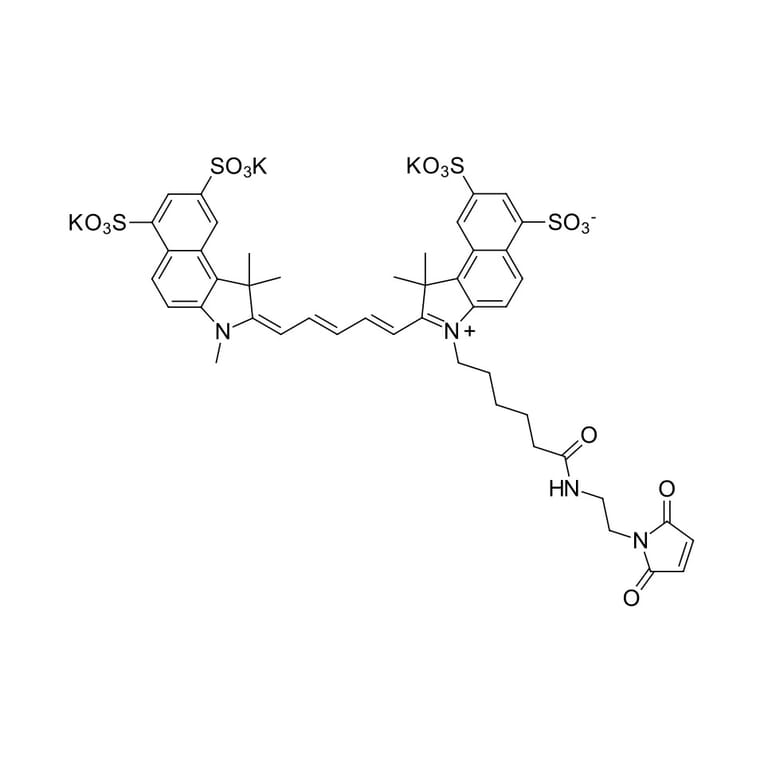
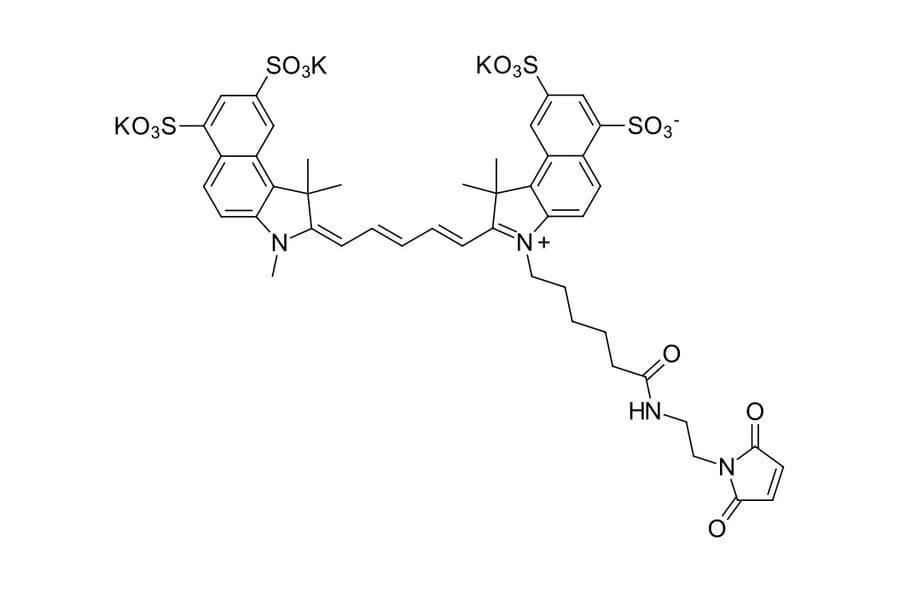
Description
Sulfo-Cyanine 5.5 dye is a water soluble far red to NIR emitting dye which is very hydrophilic due to the presence of four sulfo groups. Sulfo-Cyanine 5.5 derivatives exhibit high water solubility. The dye is a perfect choice for the labeling of sensitive proteins, nanoparticles, and highly hydrophylic biopolymers. This maleimide is a thiol reactive dye that selectively labels cysteine residues. Disulfide bonds of native proteins should be reduced with an appropriate reducing agent, such as TCEP, according to our recommended protocol.