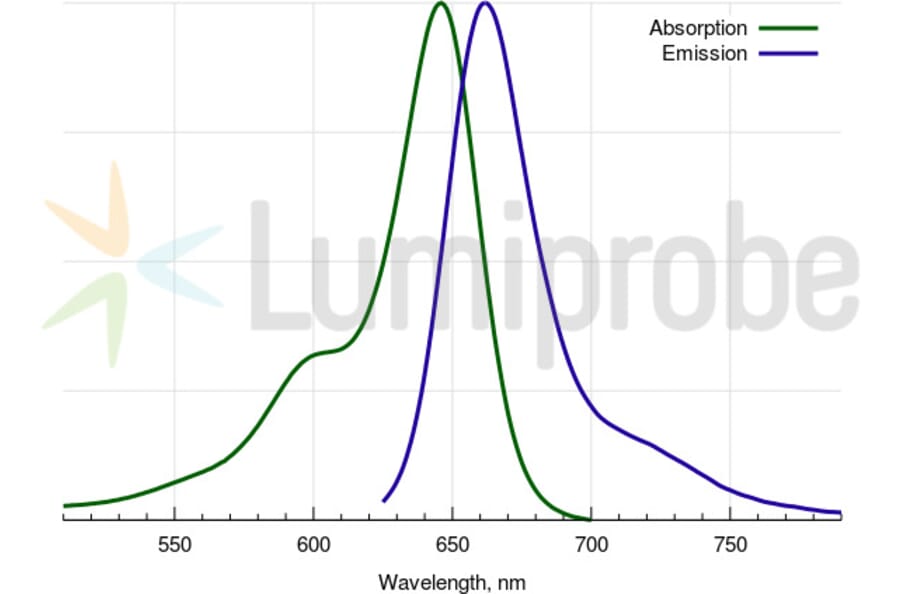
Description
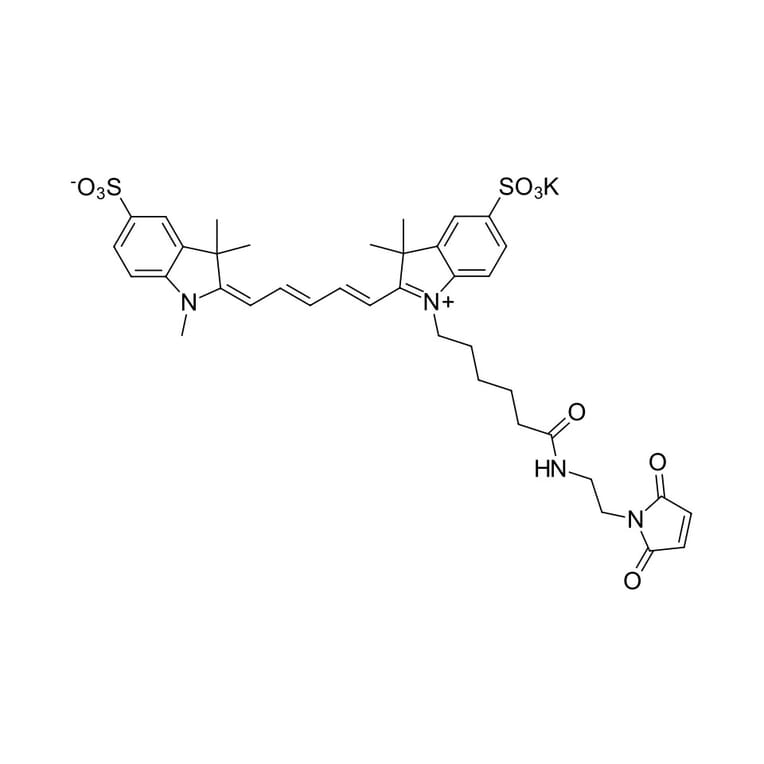
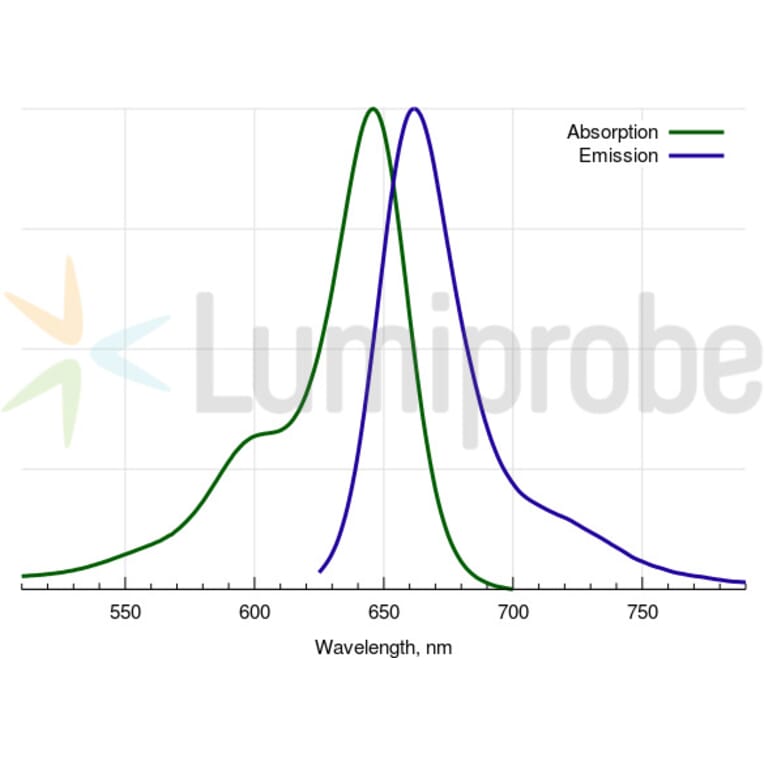
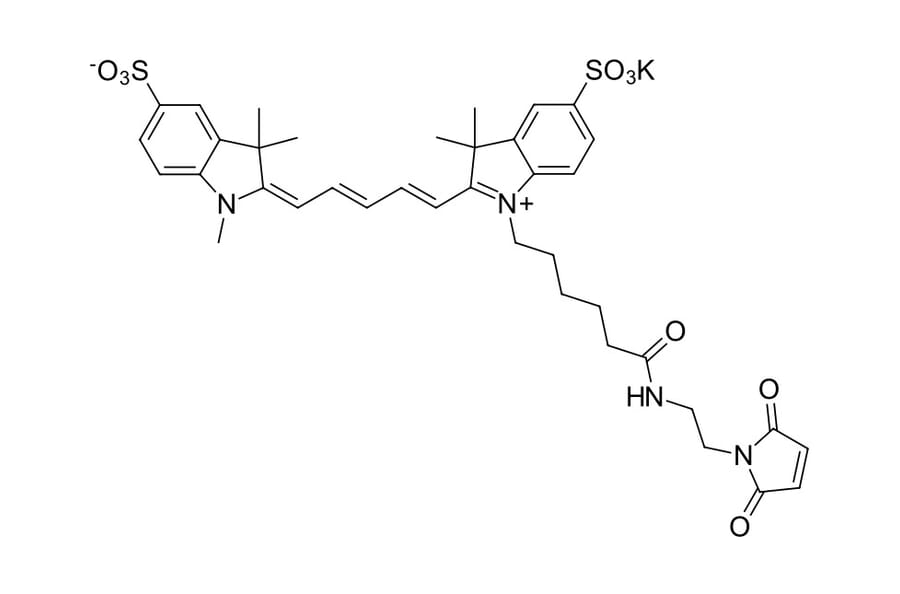
A water soluble, hydrophilic sulfo-Cyanine 5 maleimide (Cy5® maleimide analog). We recommend this product for protein labeling, including labeling of antibodies as a perfect replacement for Cy5® maleimide. Labeled proteins can be easily separated from unreacted dye by gel filtration, spin column purification, dialysis, electrophoresis or chromatography. An analog without sulfo groups, Cyanine 5 maleimide, is also available.