Primary Antibodies
Secondary Antibodies
Proteins & Peptides
ELISA Kits
About Us
Contact Us
Sign In/Register
0
ISO 9001:2015 Certified
Live Customer Support
4.5/5 on Trustpilot
100% Quality Guarantee
Home
Primary Antibodies
Rad18 Antibodies
Anti-Rad18 Antibody (A353)
Anti-Rad18 Antibody (A353)
Overview
Specifications
Images
Enlarge Image
Enlarge Image
Enlarge Image
$590
Product Datasheet
Rabbit polyclonal antibody to Rad18 for WB, IP and IF.
100% Guarantee
Price Match Guarantee
Product Size:
100µg
Quantity:
1
2
3
4
5
6
7
8
9
10
Add To Cart
Request a Quotation
Custom or Bulk Request
Shipping Information
Freight/Packing Charges:
$40
Dispatched from St. Louis, MO.
Lead Time: 4-6 business days.
Specifications
Name
Anti-Rad18 Antibody
Description
Rabbit polyclonal antibody to Rad18.
Applications
WB
,
IP
,
IF
Dilutions
WB: 1:1,000
Reactivity
Human
Immunogen
Synthetic peptide containing 113 carboxyl terminal amino acids of human Rad18 protein (GST tag).
Host
Rabbit
Clonality
Polyclonal
Isotype
IgG
Conjugate
Unconjugated
Purification
Affinity purification.
Concentration
1 mg/ml
Product Form
Liquid
Formulation
Supplied in Phosphate Buffered Saline with 50% Glycerol.
Storage
Shipped at 4°C. Upon delivery aliquot and store at -20°C. Avoid freeze / thaw cycles.
Synonyms
E3 ubiquitin-protein ligase RAD18, hHR18, hRAD18, Postreplication repair protein RAD18, RING finger protein 73, RING-type E3 ubiquitin transferase RAD18, RNF73
Isotype Controls
Rabbit IgG (A82272)
Rabbit IgG (A17360)
Suitable Secondaries
Goat Anti-Rabbit IgG H&L Antibody (AP) (A294874)
Goat Anti-Rabbit IgG H&L Antibody (Biotin) (A294795)
Goat Anti-Rabbit IgG H&L Antibody (FITC) (A294887)
Goat Anti-Rabbit IgG H&L Antibody (HRP) (A294888)
See all Anti-Rabbit IgG Secondaries →
Disclaimer
This product is for research use only. It is not intended for diagnostic or therapeutic use.
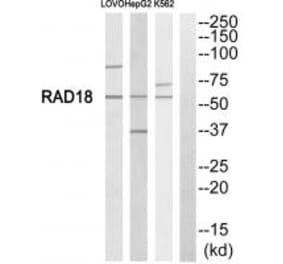
Scientific Validation Data
Validation Data
(3)
Enlarge Image
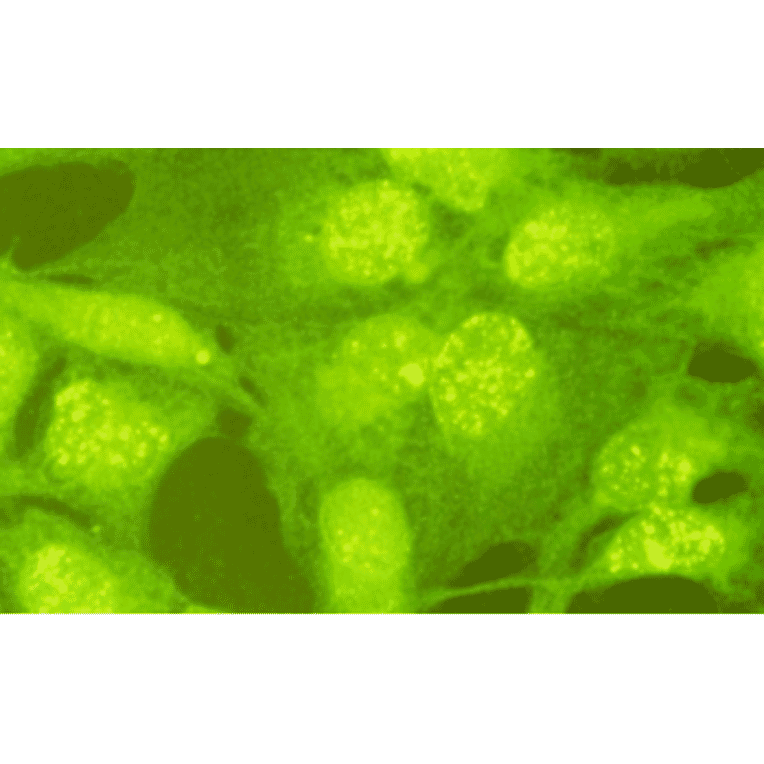
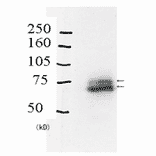
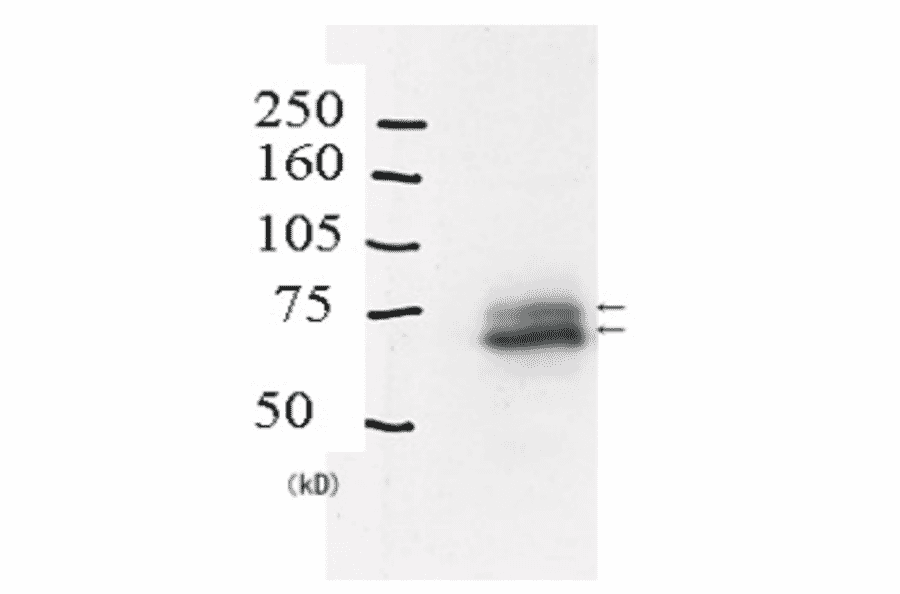
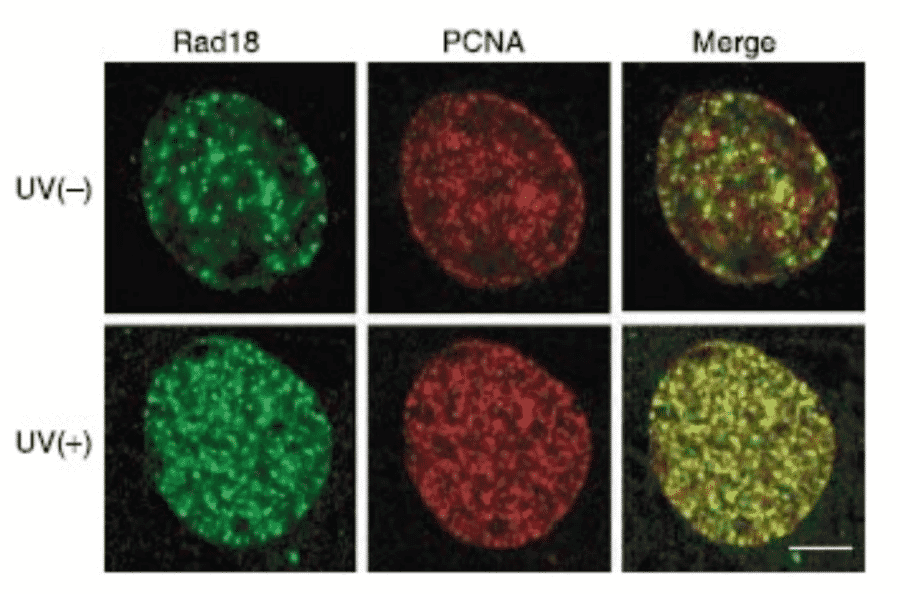
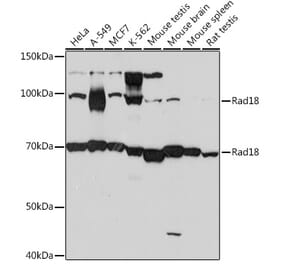
Anti-Rad18 Antibody (A353)
Anti-Rad18 Antibody
Enlarge Image
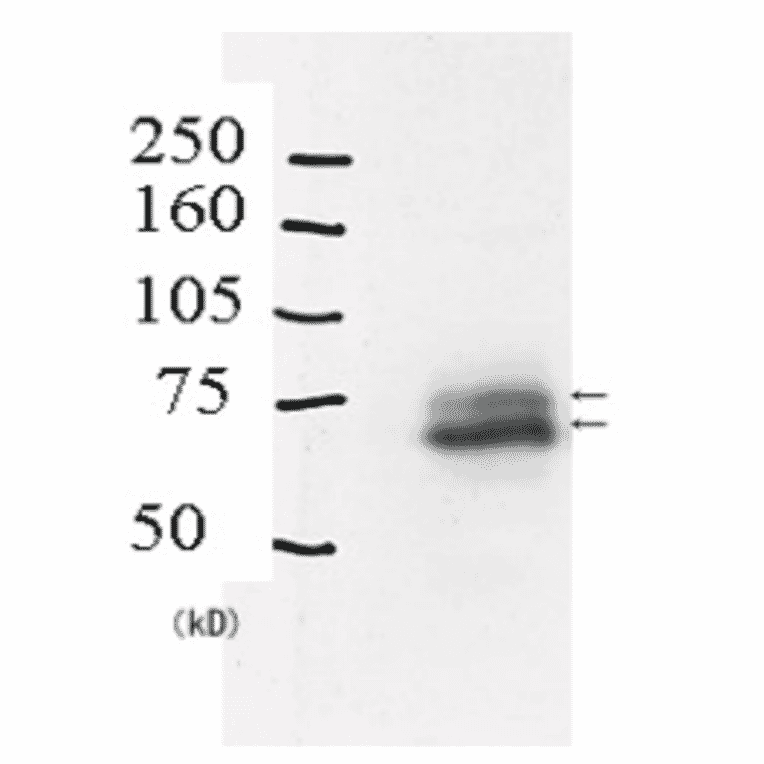
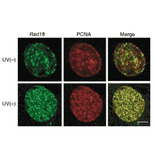
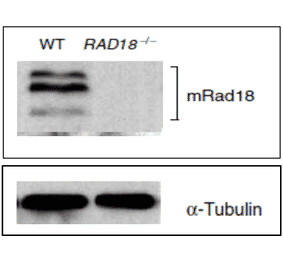
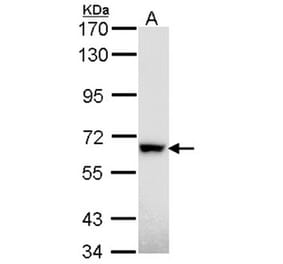
Anti-Rad18 Antibody (A353)
Anti-Rad18 Antibody
Enlarge Image
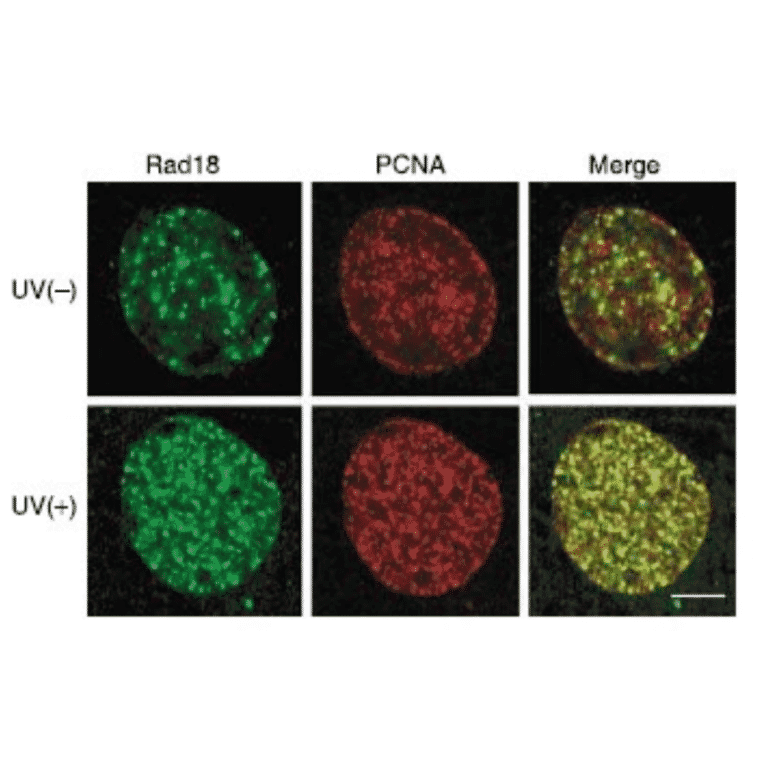
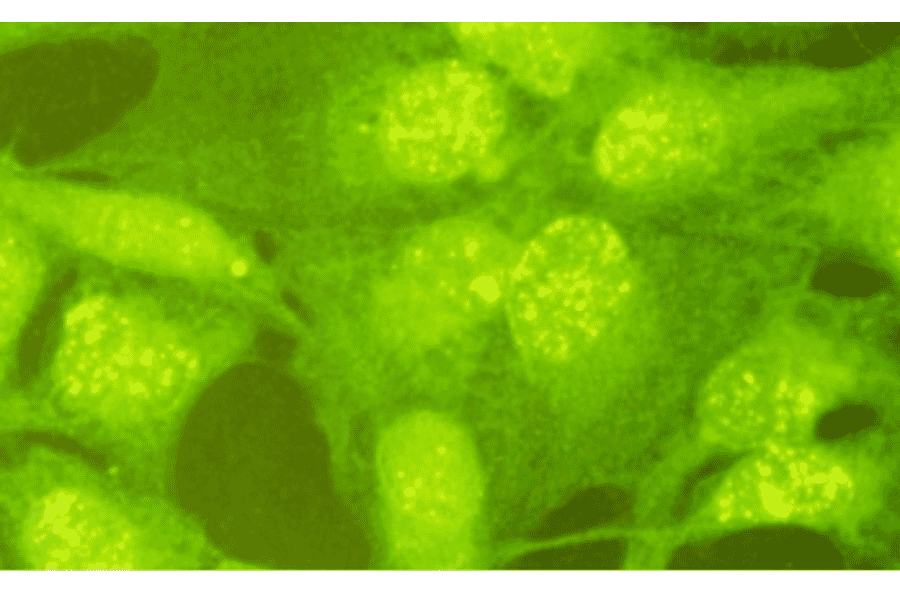
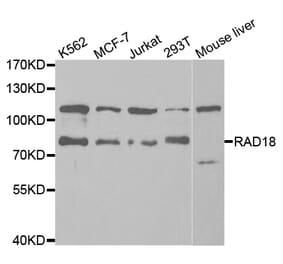
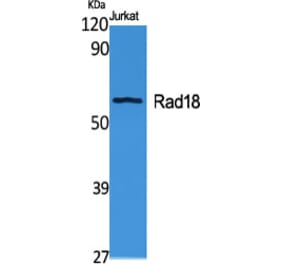
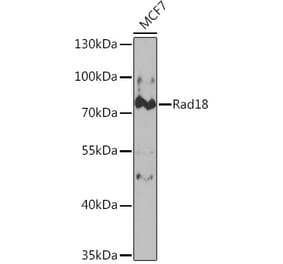
Anti-Rad18 Antibody (A353)
Anti-Rad18 Antibody
Publishing research using Anti-Rad18 Antibody (A353)? Please
let us know
so that we can list the citation on this page.
Alternative products to Anti-Rad18 Antibody (A353)
(4)
A354
Anti-Rad18 Antibody
Rabbit polyclonal antibody to Rad18 for WB, IP, IF and IHC.
(2)
A35278
Anti-RAD18 Antibody
Rabbit polyclonal antibody to RAD18 for WB, IHC and ICC/IF.
(2)
A99603
Anti-RAD18 Antibody
Rabbit polyclonal antibody to RAD18 for WB and IHC.
A348095
Anti-Rad18 Antibody [24GB880]
Recombinant rabbit monoclonal [24GB880] antibody to Rad18 for WB, Flow Cytometry and ICC/IF.
A307171
Anti-RAD18 Antibody [ARC1058]
Rabbit monoclonal [ARC1058] antibody to RAD18 for WB.
(2)
A37490
Anti-RAD18 Antibody
Rabbit polyclonal antibody to RAD18 for WB and IHC.
A14717
Anti-RAD18 Antibody
Rabbit polyclonal antibody to RAD18 for WB.
A42929
Anti-RAD18 Antibody
Rabbit polyclonal antibody to RAD18 for WB.
(4)
A354
Anti-Rad18 Antibody
Rabbit polyclonal antibody to Rad18 for WB, IP, IF and IHC.
(2)
A35278
Anti-RAD18 Antibody
Rabbit polyclonal antibody to RAD18 for WB, IHC and ICC/IF.
(2)
A99603
Anti-RAD18 Antibody
Rabbit polyclonal antibody to RAD18 for WB and IHC.
A348095
Anti-Rad18 Antibody [24GB880]
Recombinant rabbit monoclonal [24GB880] antibody to Rad18 for WB, Flow Cytometry and ICC/IF.
A307171
Anti-RAD18 Antibody [ARC1058]
Rabbit monoclonal [ARC1058] antibody to RAD18 for WB.
(2)
A37490
Anti-RAD18 Antibody
Rabbit polyclonal antibody to RAD18 for WB and IHC.
A14717
Anti-RAD18 Antibody
Rabbit polyclonal antibody to RAD18 for WB.
A42929
Anti-RAD18 Antibody
Rabbit polyclonal antibody to RAD18 for WB.
See all Rad18 Antibodies
Proteins predicted to interact with Rad18
Predicted protein interactions based upon String database. Revelancy score correlates with probability of interaction.
Rad6 Antibodies
99.9% Relevancy Score
UBE2B Antibodies
99.9% Relevancy Score
PCNA Antibodies
99.8% Relevancy Score
HLTF Antibodies
99.6% Relevancy Score
Ubiquitin Antibodies
99.5% Relevancy Score
SHPRH Antibodies
99.4% Relevancy Score
Rad51 Antibodies
99.4% Relevancy Score
MMS2 Antibodies
99.2% Relevancy Score
UBC13 Antibodies
98.8% Relevancy Score
Top