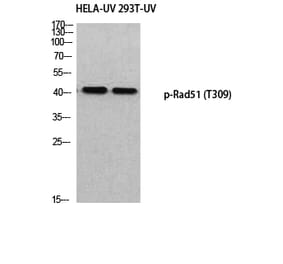
Unconjugated
The minichromosome maintenance (MCM) helicase physically interacts with the recombination proteins Rad51 and Rad52 from yeast to human cells. We show, in Saccharomyces cerevisiae, that these interactions occur within a nuclease-insoluble scaffold enriched in replication/repair factors. Rad51 accumulates in a MCM- and DNA-binding-independent manner and interacts with MCM helicases located outside of the replication origins and forks. MCM, Rad51, and Rad52 accumulate in this scaffold in G1 and are released during the S phase. In the presence of replication-blocking lesions, Cdc7 prevents their release from the scaffold, thus maintaining the interactions. We identify a rad51 mutant that is impaired in its ability to bind to MCM but not to the scaffold. This mutant is proficient in recombination but partially defective in single-stranded DNA (ssDNA) gap filling and replication fork progression through damaged DNA. Therefore, cells accumulate MCM/Rad51/Rad52 complexes at specific nuclear scaffolds in G1 to assist stressed forks through non-recombinogenic functions.
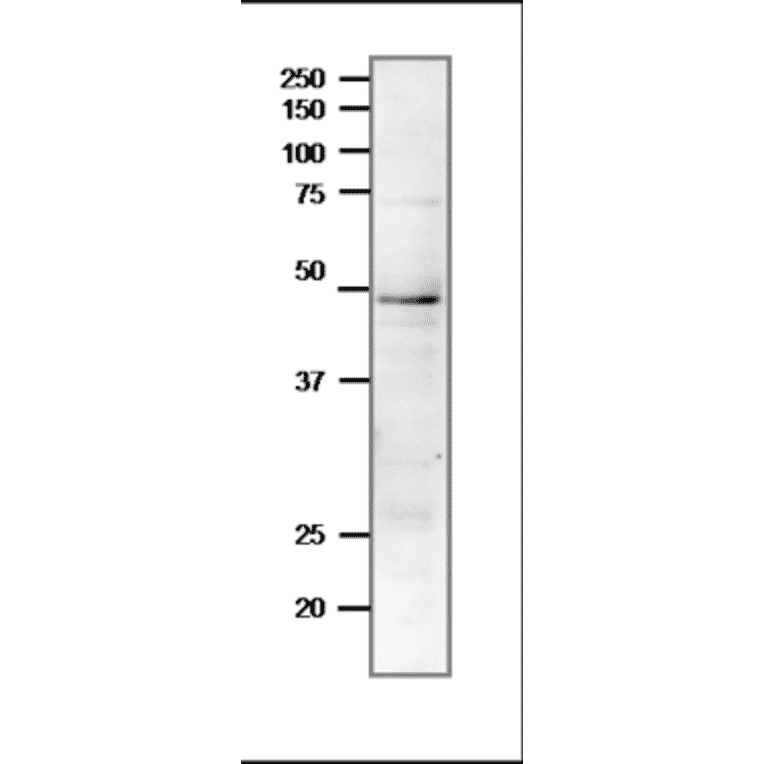
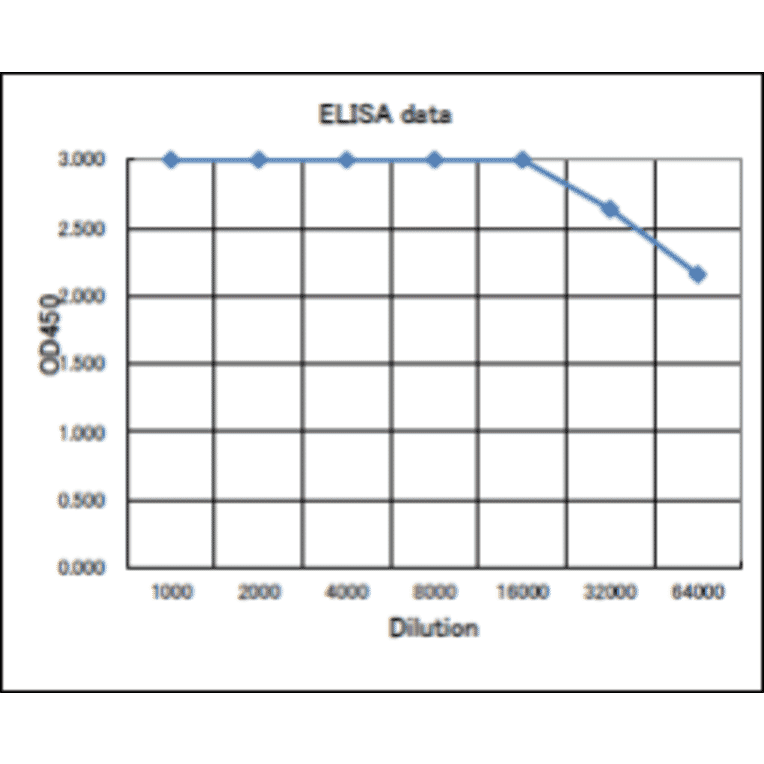

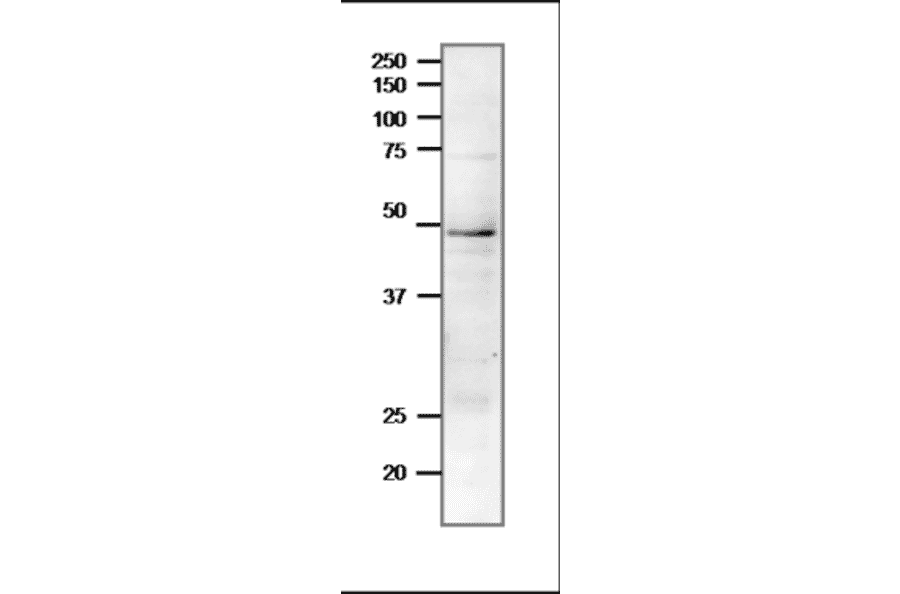
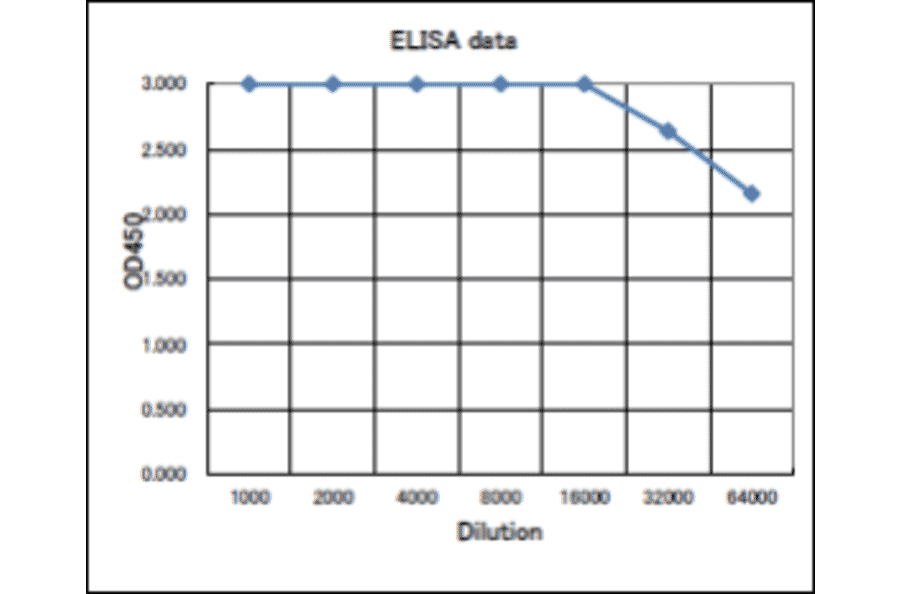

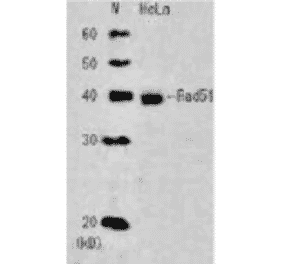
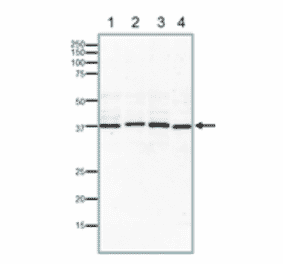

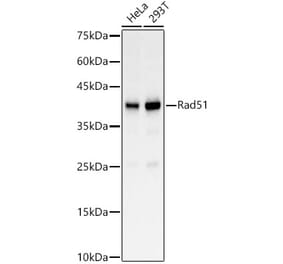

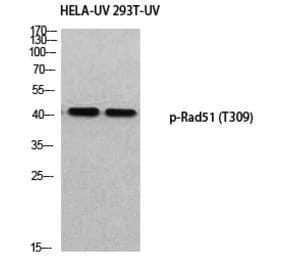
![Protein Array - Anti-RAD51 Antibody [RAD51/2701] - BSA and Azide free (A249828) - Antibodies.com](https://cdn.antibodies.com/image/catalog/253/A253008_1.jpg?profile=product_alternative)
![Protein Array - Anti-RAD51 Antibody [RAD51/2701] (A253007) - Antibodies.com](https://cdn.antibodies.com/image/catalog/249/A249828_1.jpg?profile=product_alternative)
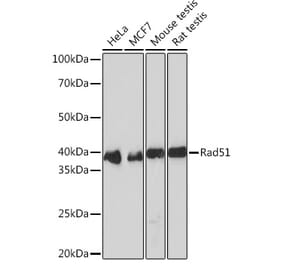
![Immunohistochemistry - Anti-RAD51 Antibody [RAD51/2702] (A253203) - Antibodies.com](https://cdn.antibodies.com/image/catalog/249/A249829_1.jpg?profile=product_alternative)
![Immunohistochemistry - Anti-RAD51 Antibody [RAD51/2702] - BSA and Azide free (A249829) - Antibodies.com](https://cdn.antibodies.com/image/catalog/253/A253009_1.jpg?profile=product_alternative)